Bacillariophyceae
pennate diatoms
David G. Mann


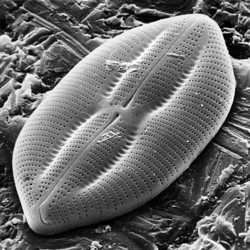


This tree diagram shows the relationships between several groups of organisms.
The root of the current tree connects the organisms featured in this tree to their containing group and the rest of the Tree of Life. The basal branching point in the tree represents the ancestor of the other groups in the tree. This ancestor diversified over time into several descendent subgroups, which are represented as internal nodes and terminal taxa to the right.

You can click on the root to travel down the Tree of Life all the way to the root of all Life, and you can click on the names of descendent subgroups to travel up the Tree of Life all the way to individual species.
For more information on ToL tree formatting, please see Interpreting the Tree or Classification. To learn more about phylogenetic trees, please visit our Phylogenetic Biology pages.
close boxIntroduction
The name Bacillariophyceae has been used in several ways: to refer to all diatoms (e.g. when the diatoms are trated as a class within the Heterokontophyta, as by van den Hoek et al. 1995), or to refer to the raphe-bearing pennate diatoms (e.g. Round et al. 1990), or to refer to all pennate diatoms (Medlin & Kaczmarska 2004). Here, we adopt the third of these: all pennate diatoms.
The pennate diatoms are characterized morphologically by the feather-like organization of their valve pattern, which is evident in the title illustrations. The distinction between this kind of pattern and the centric patterns found in other clades of diatoms was first discussed by Schütt (1896) and Karsten (1928). In pennates, a long rib or strip of plain silica runs along the length of the valve, subtending transverse ribs and lines of pores on either side. In contrast, the clades of centric diatoms possess a radial organization of the pattern. Electron microscopy has revealed that, at the centre of the radial pattern, there is a ring-like annulus (although the annulus can be elliptical or elongate in some species, such as Odontella sinensis and Attheya species: Pickett-Heaps et al. 1990, Stonik et al. 2006). Because of the resemblance of the pennate valve to the human rib-cage, the longitudinal rib or strip is called the sternum.
Most pennate diatoms have elongate cells, shaped like boats, rods, spicules or bananas and it is sometimes stated that this, rather than the organization of the valve pattern, is the essential feature of the group. However, elongate shape is not confined to pennate diatoms, being found in several clades of polar centric diatoms.
Pennate diatoms are monophyletic according to molecular phylogenies (e.g. Medlin & Kaczmarska 2004, Sorhannus 2007): the sternum has apparently evolved only once. Another autapomorphy of pennates is the loss of all flagellate stages: during sexual reproduction, fusion takes place between large amoeboid or motionless gametes, which are usually ± equal in size (morphological isogamy).
The pennates are more species-rich than other diatom groups. Most species are benthic, growing attached to solid substrata or moving through sediments and over surfaces, but the group also contains some common and important planktonic genera, such as the toxin-producing Pseudo-nitzschia, and the remarkable hair-like Thalassiothrix, which can grow up to 5 mm long.
Characteristics
- Valve pattern feather-like, organized with respect to a longitudinal sternum.
- Sexual reproduction via morphological isogamy (or slight anisogamy) of amoeboid or swelling static gametes.
- Cells mostly elongate, bipolar.
- Shape usually created through the constraining effects of a set of special silicified bands (the perizonium), formed during auxospore expansion.
- Predominantly benthic.
- Rimoportulae or their presumed homologues, the raphe slits, present in most genera and species.
Discussion of Phylogenetic Relationships
Evolution within the Bacillariophyceae is better understood than among the radial and polar centric diatoms. The earliest molecular systematic paper on diatom evolution (Medlin et al. 1993) indicated three clades:
- A clade of pennate diatoms lacking a raphe system (Rhaphoneis, Asterionellopsis)
- A second clade of pennate diatoms lacking a raphe system (Thalassionema, Fragilaria)
- Raphid pennate diatoms (Bacillaria, Nitzschia, Cylindrotheca)
The relationship among these was {1(2,3)}. In other words, the raphid diatoms are not the sister group to all of the other pennate diatoms, which lack a raphe system. Instead they are the sister group of a particular subset of the araphid pennate diatoms. At present, none of the three clades, nor the (2,3) clade, have agreed names. Clade 3 corresponds to the Bacillariophyceae sensu Round et al. (1990), but Medlin & Kaczmarska (2004), followed by Adl et al. (2005) used this name for all pennate diatoms, as here.
Phylogenetic analyses published since 1993, including many more taxa, still generally show the same {1(2,3)} structure (e.g. Kooistra et al. 2007, Sorhannus 2007). For the moment, the three clades are given informal names here. Clade I araphids comprise a small, morphologically disparate group, which are all marine. Clade II araphids comprise the majority of the Fragilariophyceae sensu Round et al. (1990), i.e. of the pennate diatoms that lack a raphe system. The sister relationship between the raphid diatoms and clade II shows that none of the subclades within clade II are likely to furnish clues as to how the raphe evolved.
The consequence of the {1(2,3)} relationship is that the pennate diatoms that lack a raphe system, which were recognized as the class Fragilariophyceae by Round et al. (1990), are a paraphyletic assemblage. Some people therefore reject any use of the phrase 'araphid pennate diatoms'.
The position of the family Striatellaceae, containing Striatella and Pseudostriatella (but not Hyalosira and Grammatophora as Round et al. 1990 thought) is currently unclear, since it varies greatly from analysis to analysis.
The earliest known pennates in the fossil record are Sceptroneis and Incisoria from the Late Cretaceous (Hajós & Stradner 1975), which appear to belong to the clade I family Rhaphoneidaceae, and Rhaphoneis itself is also reported from the Late Cretaceous (Sims et al. 2006). A few further araphid pennates occur in Cretaceous deposits but their affinities are less clear. Fossil raphid diatoms are not known until the Tertiary (Palaeocene: Sims et al. 2006), but the group probably arose slightly earlier, in the Late Cretaceous (Sorhannus 2007 estimates 75 Mya for the primary diversification of the raphids and 93.8 Ma for the separation of the raphids and clade II).
References
Hajós, M. & Stradner,H. (1975). Late Cretaceous Archaeomonadaceae, Diatomaceae, and Silicoflagellatae from the South Pacific Ocean. Deep Sea Drilling Project Leg 29, site 275. Initial Reports of the Deep Sea Drilling Project 29: 913–1109.
Karsten,G. (1928). Bacillariophyta (Diatomaceae). In: Die Natürlichen Pflanzenfamilien, 2nd ed. (Ed. A. Engler & K. Prantl), vol. 2 Leipzig, W. Engelmann, pp. 105–203.
Kooistra, W.H.C.F., Gersonde, R., Medlin, L.K. & Mann, D.G. (2007). The origin and evolution of the diatoms: their adaptation to a planktonic existence. In: Evolution of primary producers in the sea (Ed. by P.G. Falkowski & A.H. Knoll), pp. 207-249. Elsevier Academic Press, Burlington, Massachusetts.
Medlin, L.K. & Kaczmarska, I. (2004). Evolution of the diatoms: V. Morphological and cytological support for the major clades and a taxonomic revision. Phycologia 43: 245–270.
Pickett-Heaps, J.D., Schmid, A.-M.M., Edgar, L.A. (1990). the cell biology of diatom valve formation. Progress in Phycological Research 7: 1–168.
Round, F.E., Crawford, R.M. & Mann, D.G. (1990). The diatoms. Biology and morphology of the genera. Cambridge University Press, Cambridge. 747 pp.
Schütt, F. (1896). Bacillariales (Diatomeae). In: Die natürlichen Pflanzenfamilien (Engler, A. and Prantl, K., eds.), I. lb. Leipzig (1896).
Sims, P.A., Mann, D.G. & Medlin, L.K. (2006). Evolution of the diatoms: insights from fossil, biological and molecular data. Phycologia 45: 361-402.
Sorhannus, U. (2007). A nuclear-encoded small-subunit ribosomal RNA timescale for diatom evolution. Marine Micropaleontology 65: 1-12.
Stonik, I.V., Orlova, T.Yu. & Crawford, R.M. (2006). Attheya ussurensis sp. nov. (Bacillariophyta) - a new marine diatom from the coastal waters of the Sea of Japan and a reappraisal of the genus. Phycologia 45: 141-147.
van den Hoek, C., Mann, D.G., Jahns, H.M. (1995). Algae. An introduction to phycology. Cambridge University Press, Cambridge. 623 pp.
Title Illustrations

| Scientific Name | Asterionellopsis glacialis |
|---|---|
| Comments | Transmission electron micrograph of basal section of valve, with pore field |
| Creator | Frank E. Round |
| Acknowledgements | This image is derived from the Professor Frank Round Image Archive at the Royal Botanic Garden Edinburgh |
| Specimen Condition | Dead Specimen |
| Identified By | David Mann |
| Life Cycle Stage | Vegetative phase |
| Body Part | Valve |
| View | Whole mount |
| Image Use |
 This media file is licensed under the Creative Commons Attribution-NonCommercial License - Version 3.0. This media file is licensed under the Creative Commons Attribution-NonCommercial License - Version 3.0.
|
| Copyright |
© 2008 David G. Mann

|

| Scientific Name | Neidium |
|---|---|
| Location | Ashford in the Water, Derbyshire, England |
| Specimen Condition | Live Specimen |
| Identified By | David Mann |
| Life Cycle Stage | Vegetative phase |
| Body Part | Whole cell |
| View | Median optical section |
| Image Use |
This media file is licensed under the Creative Commons Attribution-NonCommercial License - Version 3.0.
|
| Copyright |
© 2008 David G. Mann
|
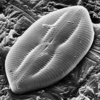
| Scientific Name | Lyrella atlantica |
|---|---|
| Specimen Condition | Dead Specimen |
| Identified By | David Mann |
| Life Cycle Stage | Vegetative phase |
| Body Part | Valve, SEM |
| View | External, tilted |
| Image Use |
This media file is licensed under the Creative Commons Attribution-NonCommercial License - Version 3.0.
|
| Copyright |
© 2008 David G. Mann
|
About This Page
This page is being developed as part of the Tree of Life Web Project Protist Diversity Workshop, co-sponsored by the Canadian Institute for Advanced Research (CIFAR) program in Integrated Microbial Biodiversity and the Tula Foundation.
David G. Mann
Royal Botanic Garden Edinburgh, United Kingdom
Correspondence regarding this page should be directed to David G. Mann at
d.mann@rbge.org.uk
Page copyright © 2008 David G. Mann
 Page: Tree of Life
Bacillariophyceae. pennate diatoms.
Authored by
David G. Mann.
The TEXT of this page is licensed under the
Creative Commons Attribution-NonCommercial License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
Page: Tree of Life
Bacillariophyceae. pennate diatoms.
Authored by
David G. Mann.
The TEXT of this page is licensed under the
Creative Commons Attribution-NonCommercial License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
- First online 07 February 2010
Citing this page:
Mann, David G. 2010. Bacillariophyceae. pennate diatoms. Version 07 February 2010 (under construction). http://tolweb.org/Bacillariophyceae/125298/2010.02.07 in The Tree of Life Web Project, http://tolweb.org/